The effects of ��-MSH on TUNEL staining and cell viability in the macrophages must be post-caspase activity in the apoptotic pathway. It has been shown that inhibiting phosphorylation of p38 MAPK inhibits the apoptosis pathway before DNA fragmentation. It is not clear if p38 MAPK AbMole Metaproterenol Sulfate activation is directly involved in the activation of DNA fragmentation, or that its activation is a bystander indicator of Caspase 3 activation of MAP kinase kinases that initiates the process of DNA fragmentation. While the 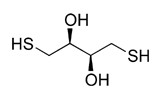 intracellular pathways of ��-MSH mediated signaling are not fully understood, it is known that ��MSH suppresses the activation of both NF-kB and p38 MAPK AbMole Terbuthylazine through its melanocortin receptors MC1r and MC3r. This suppresses inflammatory activity in the macrophages stimulated through Toll-like receptors and proinflammatory cytokines. The suppression of NF-kB is ��-MSH promoting IRAK-M binding to IRAK-1 to inhibit TLR signaling. The suppression of p38 MAPK is though ��-MSH activation of cAMP-dependent PKA. Therefore, since the results show no effect of ��-MSH on the initial steps and the caspase steps of apoptosis, and that ��-MSH is known to suppress p38 MAPK phosphorylation, it suggests that the effects of ��-MSH are on the post-caspase steps of apoptosis to promote cell survival. The survival of such affected macrophages may be part of immune-homeostasis mediated by ��-MSH within localized tissue microenvironments. We know that macrophages treated with ��-MSH are suppressed in inflammatory activity, and may promote regulatory or tolerogenic immunity. In the retina, ��-MSH is part of RPE mediated induction of suppressor macrophages and similar characteristics in the retinal microglial cells. However, the signaling pathways that result in activation of genes and functions of inflammation within macrophages are also signals of survival and resistance to apoptosis. Therefore, in the healthy retina ��-MSH must be providing an anti-inflammatory signal, and a necessary survival signal for the immunosuppressive microglial to function and survive in the retina. Moreover, ��-MSH may be part of a selective mechanism mediated by the RPE to exclude macrophages that are unresponsive to ��-MSH immunosuppressive activity. In this way macrophages and possibly dendritic cells migrating into the retina would encounter the soluble apoptotic signal from RPE and will have to respond to ��-MSH to survive, but by doing so makes them immunosuppressive. This selective process could explain why in the healthy retina the microglial cells are found to be characteristically uniform as myeloid suppressor cell-like. Some of the caspases also have non-apoptotic activities. The diversion of the caspases to other non-apoptotic activities could be the effects of ��-MSH with saving the macrophages from apoptosis. A non-apoptotic activity of Caspase 8 is to activate NF-��B. The suppression of Caspase 8 that was detected could be ��-MSH suppressing another pathway to NF-��B activation. Maintaining nonapoptotic activities of Caspase 3 has benefits to ocular immunobiology. The non-apoptotic functions of Caspase 3 include suppression of MHC class II antigen expression and dendritic cell maturation. This would greatly contribute to the immune privileged and immunosuppressive microenvironment of the retina. Since ��-MSH can have a role in immune-homeostasis to resolve inflammation, it could mediate more than immunosuppression by maintaining in surviving cells caspase functionalities that contribute to resolving inflammation and promoting wound repair. The neuropeptide ��-MSH is not just an anti-inflammatory factor. It modulates the functionality of immune cells.
intracellular pathways of ��-MSH mediated signaling are not fully understood, it is known that ��MSH suppresses the activation of both NF-kB and p38 MAPK AbMole Terbuthylazine through its melanocortin receptors MC1r and MC3r. This suppresses inflammatory activity in the macrophages stimulated through Toll-like receptors and proinflammatory cytokines. The suppression of NF-kB is ��-MSH promoting IRAK-M binding to IRAK-1 to inhibit TLR signaling. The suppression of p38 MAPK is though ��-MSH activation of cAMP-dependent PKA. Therefore, since the results show no effect of ��-MSH on the initial steps and the caspase steps of apoptosis, and that ��-MSH is known to suppress p38 MAPK phosphorylation, it suggests that the effects of ��-MSH are on the post-caspase steps of apoptosis to promote cell survival. The survival of such affected macrophages may be part of immune-homeostasis mediated by ��-MSH within localized tissue microenvironments. We know that macrophages treated with ��-MSH are suppressed in inflammatory activity, and may promote regulatory or tolerogenic immunity. In the retina, ��-MSH is part of RPE mediated induction of suppressor macrophages and similar characteristics in the retinal microglial cells. However, the signaling pathways that result in activation of genes and functions of inflammation within macrophages are also signals of survival and resistance to apoptosis. Therefore, in the healthy retina ��-MSH must be providing an anti-inflammatory signal, and a necessary survival signal for the immunosuppressive microglial to function and survive in the retina. Moreover, ��-MSH may be part of a selective mechanism mediated by the RPE to exclude macrophages that are unresponsive to ��-MSH immunosuppressive activity. In this way macrophages and possibly dendritic cells migrating into the retina would encounter the soluble apoptotic signal from RPE and will have to respond to ��-MSH to survive, but by doing so makes them immunosuppressive. This selective process could explain why in the healthy retina the microglial cells are found to be characteristically uniform as myeloid suppressor cell-like. Some of the caspases also have non-apoptotic activities. The diversion of the caspases to other non-apoptotic activities could be the effects of ��-MSH with saving the macrophages from apoptosis. A non-apoptotic activity of Caspase 8 is to activate NF-��B. The suppression of Caspase 8 that was detected could be ��-MSH suppressing another pathway to NF-��B activation. Maintaining nonapoptotic activities of Caspase 3 has benefits to ocular immunobiology. The non-apoptotic functions of Caspase 3 include suppression of MHC class II antigen expression and dendritic cell maturation. This would greatly contribute to the immune privileged and immunosuppressive microenvironment of the retina. Since ��-MSH can have a role in immune-homeostasis to resolve inflammation, it could mediate more than immunosuppression by maintaining in surviving cells caspase functionalities that contribute to resolving inflammation and promoting wound repair. The neuropeptide ��-MSH is not just an anti-inflammatory factor. It modulates the functionality of immune cells.
It promoting alternative activation of macrophages and dendritic cells in that they suppress inflammation and activate
Leave a reply
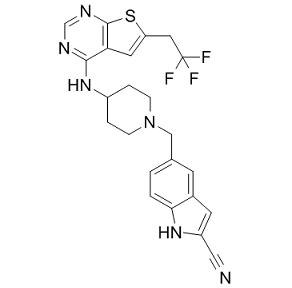 rabbit, although intimal thickening and medial calcification were seen. This result indicated that periaortic incubation of 0.5 mol/L CaCl2 for 20 min causes an increase in wall thickness with extensive intimal hyperplasia and calcification, but no formation of aneurysm. Freestone et al. reported the same findings, there was no aneurysmal dilatation of the aorta after periaortic application of 0.5 mol/L CaCl2, and rapid aneurysmal dilatation only occurred after periaortic application of CaCl2 and thioglycollate in cholesterol-fed rabbits. However, the SMC, elastin and type III collagen content decreased significantly in Group C, indicating that periaortic CaCl2 incubation might promote aneurysm formation and progression when combined with elastase incubation. Of note, there was no atherosclerosis in our model, and a cholesterol-enriched diet was important in CaCl2-induced AAA formation. We did not perform any pharmacologic manipulation, such as doxycycline, propranolol or simvastatin with this model to demonstrate an impact in aneurysm formation. We will study the influence of pharmacologic intervention on aneurysm formation and development in our novel model in the future. In conclusion, we present an easy, efficient and reproducible way to create rapid dilation of rabbit aortic arteries to form a model of AAA. This simpler aneurysm model could be valuable for elucidating AAA mechanisms and therapeutic interventions, especially through drug-eluting stents or stent graft-mediated gene delivery system in experimental studies.
rabbit, although intimal thickening and medial calcification were seen. This result indicated that periaortic incubation of 0.5 mol/L CaCl2 for 20 min causes an increase in wall thickness with extensive intimal hyperplasia and calcification, but no formation of aneurysm. Freestone et al. reported the same findings, there was no aneurysmal dilatation of the aorta after periaortic application of 0.5 mol/L CaCl2, and rapid aneurysmal dilatation only occurred after periaortic application of CaCl2 and thioglycollate in cholesterol-fed rabbits. However, the SMC, elastin and type III collagen content decreased significantly in Group C, indicating that periaortic CaCl2 incubation might promote aneurysm formation and progression when combined with elastase incubation. Of note, there was no atherosclerosis in our model, and a cholesterol-enriched diet was important in CaCl2-induced AAA formation. We did not perform any pharmacologic manipulation, such as doxycycline, propranolol or simvastatin with this model to demonstrate an impact in aneurysm formation. We will study the influence of pharmacologic intervention on aneurysm formation and development in our novel model in the future. In conclusion, we present an easy, efficient and reproducible way to create rapid dilation of rabbit aortic arteries to form a model of AAA. This simpler aneurysm model could be valuable for elucidating AAA mechanisms and therapeutic interventions, especially through drug-eluting stents or stent graft-mediated gene delivery system in experimental studies.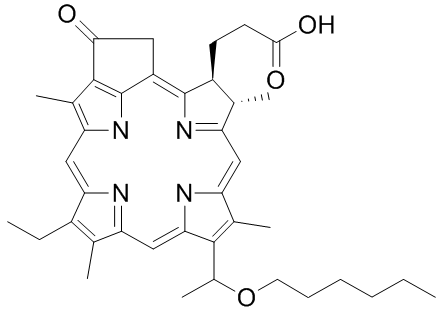 mechanisms of feedback between them. While high-throughput measurements have examined both independently in bulk samples, no combined cellto-cell analysis of these two critical components of immune response has yet been performed. We began our antibody sequence analysis by focusing on the antibody heavy chain, both because it bears the greatest responsibility for antibody specificity in general and because its large VDJ-junctional region avails it of far greater diversity as compared to the light chain. Moreover, this extended junctional diversity, generated randomly from somatic gene recombination undergone during B-cell development, provides crucial information about the common ancestry of B-cells with the same V/J recombination. This information can be used to illuminate the processes of B-cell development and clonal expansion in data-sets of arbitrary size. We classified antibody sequences according to clonal lineage by performing single-linkage clustering on their heavy chain CDR3-regions, with a distance-threshold of one amino acid, and found evidence for clonal expansion exclusively among PE+ B-cells. This helped to confirm that no systematic cross-contamination had occurred between cell samples. Using these heavy-chain clonal lineage designations as a benchmark, light-chains that had been successfully sequenced were filtered according to a statistical test that quantified the likelihood of a heavy/light-chain pairing to have occurred randomly, in order to eliminate potential PCR-contaminants. This further helped correct for B cell clonal lineages that had been mis-assigned as distinct from one another based on heavy chain sequence alone. We also analyzed our raw data directly, without using light-chain sequence-filtering or correction of antibody clonal lineages, and observed qualitatively identical results. The B-cell lineages originated in several distinct V/J-combinations. However, no overlap among clonal lineages, as characterized by their heavychain CDR3 regions, was found between PE+ and PEpopulations. We next analyzed antibody gene mutational content by tallying synonymous and non-synonymous mutations for each cell’s heavy and light chains. The isotype correlation and anti-correlation with somatic mutation were consistent with the standard model for B cell maturation. IgG expression, requiring AID-induced isotype classswitching, correlated positively with somatic mutation. Conversely, IgM and IgD correlated negatively with somatic mutation, affirming their preferential expression by un-activated, nonmutating B-cells. Such negative correlations, found in both mice, existed for only a few other genes. The most prominent of these were EBI-2, expressed on plasma and non-germinal center B-cells,, and the FCER2A receptor, responsible for the membrane-display of IgE antibodies on B-cells.
mechanisms of feedback between them. While high-throughput measurements have examined both independently in bulk samples, no combined cellto-cell analysis of these two critical components of immune response has yet been performed. We began our antibody sequence analysis by focusing on the antibody heavy chain, both because it bears the greatest responsibility for antibody specificity in general and because its large VDJ-junctional region avails it of far greater diversity as compared to the light chain. Moreover, this extended junctional diversity, generated randomly from somatic gene recombination undergone during B-cell development, provides crucial information about the common ancestry of B-cells with the same V/J recombination. This information can be used to illuminate the processes of B-cell development and clonal expansion in data-sets of arbitrary size. We classified antibody sequences according to clonal lineage by performing single-linkage clustering on their heavy chain CDR3-regions, with a distance-threshold of one amino acid, and found evidence for clonal expansion exclusively among PE+ B-cells. This helped to confirm that no systematic cross-contamination had occurred between cell samples. Using these heavy-chain clonal lineage designations as a benchmark, light-chains that had been successfully sequenced were filtered according to a statistical test that quantified the likelihood of a heavy/light-chain pairing to have occurred randomly, in order to eliminate potential PCR-contaminants. This further helped correct for B cell clonal lineages that had been mis-assigned as distinct from one another based on heavy chain sequence alone. We also analyzed our raw data directly, without using light-chain sequence-filtering or correction of antibody clonal lineages, and observed qualitatively identical results. The B-cell lineages originated in several distinct V/J-combinations. However, no overlap among clonal lineages, as characterized by their heavychain CDR3 regions, was found between PE+ and PEpopulations. We next analyzed antibody gene mutational content by tallying synonymous and non-synonymous mutations for each cell’s heavy and light chains. The isotype correlation and anti-correlation with somatic mutation were consistent with the standard model for B cell maturation. IgG expression, requiring AID-induced isotype classswitching, correlated positively with somatic mutation. Conversely, IgM and IgD correlated negatively with somatic mutation, affirming their preferential expression by un-activated, nonmutating B-cells. Such negative correlations, found in both mice, existed for only a few other genes. The most prominent of these were EBI-2, expressed on plasma and non-germinal center B-cells,, and the FCER2A receptor, responsible for the membrane-display of IgE antibodies on B-cells.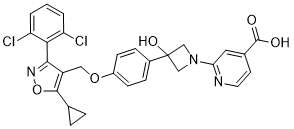 We hypothesized that one gene or a few genes within the QTL region regulate spontaneous arthritis. Accordingly, if a genomic fragment that contains the gene/s responsible for regulation of IL-1RA and development of spontaneous arthritis in BALB/c was replaced by the analogous fragment from DBA/1 mice which are resistant to spontaneous arthritis, then disease in the new strain would be reduced in incidence and/or severity. To test our hypothesis, we have developed congenic strains that contain the genomic fragments in the region of the QTL from DBA/1 mice on a BALB/c background. Congenic strains are animals in which a genetic locus has been moved from one strain/ line to the background of another strain/line by back-crossing. Polymorphism of molecular markers is used to detect the source strain of the genome components of a congenic strain. For these experiments, we used speed or marker-assisted congenic breeding. Theoretically, the classical protocol of congenic breeding needs about 10 generations, at which 99+% of the genetic background of the progeny is of recipient origin while still retaining heterozygosity at the region of interest. However, the availability of dense genetic maps of the mouse genome has allowed the development of marker-assisted breeding strategies, which reduce the number of generations required to eliminate donor strain-derived alleles outside the genetic region of interest.
We hypothesized that one gene or a few genes within the QTL region regulate spontaneous arthritis. Accordingly, if a genomic fragment that contains the gene/s responsible for regulation of IL-1RA and development of spontaneous arthritis in BALB/c was replaced by the analogous fragment from DBA/1 mice which are resistant to spontaneous arthritis, then disease in the new strain would be reduced in incidence and/or severity. To test our hypothesis, we have developed congenic strains that contain the genomic fragments in the region of the QTL from DBA/1 mice on a BALB/c background. Congenic strains are animals in which a genetic locus has been moved from one strain/ line to the background of another strain/line by back-crossing. Polymorphism of molecular markers is used to detect the source strain of the genome components of a congenic strain. For these experiments, we used speed or marker-assisted congenic breeding. Theoretically, the classical protocol of congenic breeding needs about 10 generations, at which 99+% of the genetic background of the progeny is of recipient origin while still retaining heterozygosity at the region of interest. However, the availability of dense genetic maps of the mouse genome has allowed the development of marker-assisted breeding strategies, which reduce the number of generations required to eliminate donor strain-derived alleles outside the genetic region of interest. response to EBI-2 expression, and meanwhile, EBI-2 expression is actively down-regulated among germinal center B-cells, the B-cells most actively undergoing hyper-proliferation and hypermutation. The anti-correlation observed between FCER2A expression and somatic mutation is therefore able to emerge from an indirect, but distinctly negative, relationship between the two. The correlations so far discussed were calculated across individual cells. However, one could imagine that clonal relationships might bias results, especially if phenotypic state is inherited during cell division. In order to eliminate this possibility, we used our knowledge of the B-cell population’s clonal relationships from their antibody sequences. By iteratively and randomly sampling cells from each clonal lineage, we ensured each would be equally represented in the final, averaged, correlation calculation. The resulting correlation values were qualitatively the same as those performed across cells individually. Clonal relatedness did not therefore play a significant part in defining the observed gene expressionmutation correlations. These relationships were therefore found to represent independently-sampled gene expression programs. Iterative, independent sampling of cells across each clonal antibody lineage allowed us to calculate Spearman rank-permutation p-values across the data-set for each mutation/geneexpression relationship. Broad significance was observed among the genes correlating most positively and most negatively with somatic mutation. Its consistently significant correlation with mutations may suggest an active strengthening of a B-cell’s ability to check the efficacy of mutagenesis that is coincident with the accumulation of the mutations themselves. This suggested their regulation was among the most strongly coupled to antibody proteindiversification. Surprisingly, however, the magnitudes of the differences between non-synonymous and synonymous mutation/gene-expression correlations were very different between the two mice.This stood in sharp contrast to the similarity between the magnitudes of mutation/gene-expression correlations themselves. The adaptive immune system selects B-cells that produce highspecificity antibodies to target a wide range of pathogens. Active feedback between induced antibody mutation and its effect on antibody specificity is therefore at the core of a successful immune response. In this paper, we measured this feedback by performing simultaneous measurement of gene-expression and antibody-gene variation across ensembles of individual B-cells, thereby quantifying the co-variance between these two sets of variables.
response to EBI-2 expression, and meanwhile, EBI-2 expression is actively down-regulated among germinal center B-cells, the B-cells most actively undergoing hyper-proliferation and hypermutation. The anti-correlation observed between FCER2A expression and somatic mutation is therefore able to emerge from an indirect, but distinctly negative, relationship between the two. The correlations so far discussed were calculated across individual cells. However, one could imagine that clonal relationships might bias results, especially if phenotypic state is inherited during cell division. In order to eliminate this possibility, we used our knowledge of the B-cell population’s clonal relationships from their antibody sequences. By iteratively and randomly sampling cells from each clonal lineage, we ensured each would be equally represented in the final, averaged, correlation calculation. The resulting correlation values were qualitatively the same as those performed across cells individually. Clonal relatedness did not therefore play a significant part in defining the observed gene expressionmutation correlations. These relationships were therefore found to represent independently-sampled gene expression programs. Iterative, independent sampling of cells across each clonal antibody lineage allowed us to calculate Spearman rank-permutation p-values across the data-set for each mutation/geneexpression relationship. Broad significance was observed among the genes correlating most positively and most negatively with somatic mutation. Its consistently significant correlation with mutations may suggest an active strengthening of a B-cell’s ability to check the efficacy of mutagenesis that is coincident with the accumulation of the mutations themselves. This suggested their regulation was among the most strongly coupled to antibody proteindiversification. Surprisingly, however, the magnitudes of the differences between non-synonymous and synonymous mutation/gene-expression correlations were very different between the two mice.This stood in sharp contrast to the similarity between the magnitudes of mutation/gene-expression correlations themselves. The adaptive immune system selects B-cells that produce highspecificity antibodies to target a wide range of pathogens. Active feedback between induced antibody mutation and its effect on antibody specificity is therefore at the core of a successful immune response. In this paper, we measured this feedback by performing simultaneous measurement of gene-expression and antibody-gene variation across ensembles of individual B-cells, thereby quantifying the co-variance between these two sets of variables.