To detect possible enzymatic activity-independent Danshensu effects of the phosphatase, we also generated animals transgenic for the inactive C227S mutants of LYP-W620. In the presence of LYP-W620, we observed increased phosphatase activity of LYPW620 in thymocytes, associated with decreased TCR signaling in total thymocytes and DP thymocytes. No anomalies in thymic TCR signaling were detected in animals carrying the inactive form of LYP-W620, despite supraphysiological expression of LYP directed by the transgene in these animals. Results with LYPW620 C227S variant-bearing mice thus suggest that effects of phosphatase overexpression on thymocyte TCR signaling critically depend upon enzymatic function of LYP. Overexpression of active LYP-W620 did not lead to impaired positive or Dexrazoxane hydrochloride negative selection of HY-TCR transgenic thymocytes in male mice or in OT-1 TCR-transgenic thymocytes crossed to Rip-mOva mice. Indeed, the finding of modestly-reduced autoreactive thymocyte numbers in BACLYP transgenic H-Y males suggests that LYP-W620 may actually promote negative selection of CD8 + T cells. Such a phenotype is in line with the report by Dai et al., in which PepW619-knockin autoreactive thymocytes exhibit augmented clonal deletion. We speculate that such subtle positive effects of LYP-W620 on thymocyte selection are obliterated in our TgLYP models by the combination of supraphysiological expression levels and the use of an exogenous promoter. Next, we considered the hypothesis that overexpression of the LYP-W620 variant would only impact negative selection at an intermediate TCR affinity range and thus might particularly impact those clones bearing TCR with affinity for self-antigen close to the threshold of negative selection. We addressed this hypothesis by comparing the affinity of the bulk population of Kb/ Ova-reactive T cells that survive negative selection in Rip-mOva mice. Normally, in such mice only T cells with low affinity for Kb/ Ova can be detected in the periphery. We reasoned that if LYP-W620 selectively reduces negative selection of T cells bearing antigen receptors with affinity at or just above the threshold of negative selection, then expression of TgLYPW would confer increased in the overall functional avidity of Kb/Ova-reactive T cells in Rip-mOva or Rip-mOvaxVb5 mice. Our negative data clearly argue against this hypothesis. We also considered whether LYP-W620 might selectively impact CD4 + T cell selection. The pathogenesis of disease in Skg mice depends on reduced TCR signaling in thymocytes, which results in reduced positive and negative selection of CD4 + thymocytes, but the repertoire of Skg thymic emigrants contains T cells with significant auto-reactive potential. When we overexpressed LYP-W620 in the thymus of Skg mice, we did not observe alterations of CD4 + thymocyte repertoire or a gain in disease severity. Higher affinity interactions with thymocyte TCR by autoantigens are thought to be required for the differentiation of Treg in the thymus, thus suggesting that alterations of TCR signaling in the thymus might shape the amount and repertoire of Treg. However, we did not see changes in the output of regulatory T cells from the TgLYPW Skg thymus, further arguing against the notion that LYP-W620 selectively affects high-affinity TCR interactions important for development. Importantly, in the TgLYPW model, expression of the transgene is controlled by the Lck proximal promoter and is only expressed in the thymus but not in the periphery. Therefore, we can exclude that we created a balanced situation such that 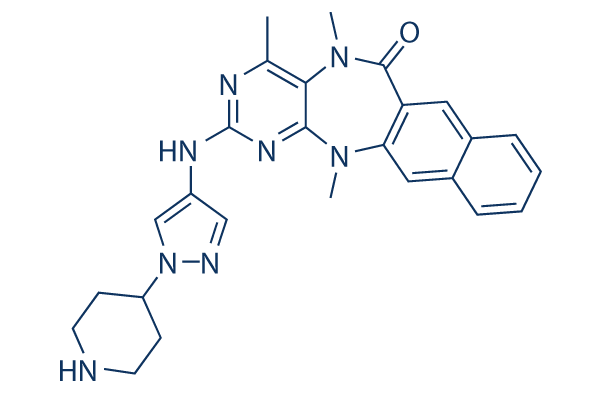 we might have augmented the thymic threshold but, at the same time, caused similar increases in the peripheral T cell activation threshold.
we might have augmented the thymic threshold but, at the same time, caused similar increases in the peripheral T cell activation threshold.
Whether the gain-of-function nature of the R620W variant is fully replicated by the R619W mutation of mouse Pep
Leave a reply
 as host cells. Host cell death is commonly induced during an infection process. The hypersensitive response, a typical ETI during which host R proteins recognize cognate pathogen effectors, is characterized by massive cell death in the local infected region to quickly deprive pathogens of water and nutrients and thereby to kill the pathogens. Although many prior studies have tested accumulation of SA and ROS and the induction of cell death as part of defense phenotype assays of pathogen-challenged plants, how these signaling molecules and cell death formation change at different time points during PTI, ETS, and ETI, has not been compared under the same experimental condition. Similar to global gene expression profiling, a detailed analysis of the behavior of these defense signaling molecules and cell death formation in plants upon pathogen attack should contribute to a better understanding of PTI, ETS, and ETI, thereby host defense mechanisms. In this report, we carefully examined several defense related phenotypes in a time course during PTI, ETS, and ETI, using Arabidopsis-P. syringae as a model system. We found that there are dynamic differences between PTI, ETS, and ETI in SA accumulation, expression of the defense marker gene PR1, and cell death formation. Such differences are dependent on the doses of the strains used. In addition, our data
as host cells. Host cell death is commonly induced during an infection process. The hypersensitive response, a typical ETI during which host R proteins recognize cognate pathogen effectors, is characterized by massive cell death in the local infected region to quickly deprive pathogens of water and nutrients and thereby to kill the pathogens. Although many prior studies have tested accumulation of SA and ROS and the induction of cell death as part of defense phenotype assays of pathogen-challenged plants, how these signaling molecules and cell death formation change at different time points during PTI, ETS, and ETI, has not been compared under the same experimental condition. Similar to global gene expression profiling, a detailed analysis of the behavior of these defense signaling molecules and cell death formation in plants upon pathogen attack should contribute to a better understanding of PTI, ETS, and ETI, thereby host defense mechanisms. In this report, we carefully examined several defense related phenotypes in a time course during PTI, ETS, and ETI, using Arabidopsis-P. syringae as a model system. We found that there are dynamic differences between PTI, ETS, and ETI in SA accumulation, expression of the defense marker gene PR1, and cell death formation. Such differences are dependent on the doses of the strains used. In addition, our data  on active MS lesions. In both studies, active lesions were defined as lesions with active demyelination as well as inflammatory infiltrates. Regarding the animal models, the original inclusion criteria were fulfilled by a study investigating three phases in MOG-induced EAE in Dark Agouti rats. In this study, female Dark Agouti rats were immunized with recombinant MOG, emulsified in complete Freund’s adjuvant. The acute phase of EAE was defined as the first EAE attack with clinical signs such as hind limb paralysis, whereas the recovery phase was defined as the first day at which the rats began to gain weight again. The relapsing phase presented as a second worsening of the clinical signs.
on active MS lesions. In both studies, active lesions were defined as lesions with active demyelination as well as inflammatory infiltrates. Regarding the animal models, the original inclusion criteria were fulfilled by a study investigating three phases in MOG-induced EAE in Dark Agouti rats. In this study, female Dark Agouti rats were immunized with recombinant MOG, emulsified in complete Freund’s adjuvant. The acute phase of EAE was defined as the first EAE attack with clinical signs such as hind limb paralysis, whereas the recovery phase was defined as the first day at which the rats began to gain weight again. The relapsing phase presented as a second worsening of the clinical signs.-chemical-structure.gif) lung findings at Ch+5 were chance occurrences among few ferrets poorly-representative of the full group experience. However, animals in both groups were randomly and blindly selected for Ch+5 sacrifice, the comparison of baseline and Ch+5 characteristics showed no significant within-group differences according to scheduled endpoint, and Ch+5 lung findings in vaccinated animals were consistent with overall clinical patterns. Nevertheless, future experiments should be powered with more animals to specifically examine these early acute clinical, immunologic, and pathologic findings and explore their possible mechanisms in greater detail. We assessed only influenza-na? ��ve animals whereas most humans, other than young children, will have prior potentially cross-attenuating influenza infection history. The use of influenzana? ��ve animals in previous swine and the current ferret studies may be relevant to the increased severity highlighted in vaccinated animals but to a lesser extent noted with the association in
lung findings at Ch+5 were chance occurrences among few ferrets poorly-representative of the full group experience. However, animals in both groups were randomly and blindly selected for Ch+5 sacrifice, the comparison of baseline and Ch+5 characteristics showed no significant within-group differences according to scheduled endpoint, and Ch+5 lung findings in vaccinated animals were consistent with overall clinical patterns. Nevertheless, future experiments should be powered with more animals to specifically examine these early acute clinical, immunologic, and pathologic findings and explore their possible mechanisms in greater detail. We assessed only influenza-na? ��ve animals whereas most humans, other than young children, will have prior potentially cross-attenuating influenza infection history. The use of influenzana? ��ve animals in previous swine and the current ferret studies may be relevant to the increased severity highlighted in vaccinated animals but to a lesser extent noted with the association in