In addition to deregulating Rho GTPases and their effectors, KAI1/CD82 may also affect the trafficking of actin-binding proteins that are associated with TEMs to alter actin organization, based on the early observations that KAI1/CD82 proteins traffic between the plasma membrane and endosomes/lysosmes and regulate the trafficking of their associated integrins. As described 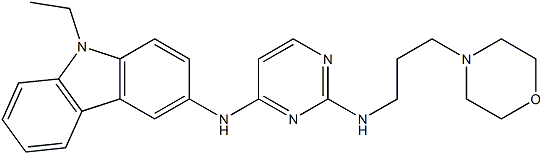 earlier, KAI1/CD82 can initiate outside-in signaling. KAI1/CD82-initiated Mepiroxol signaling may intercept the promigration signaling derived from integrin and growth factor receptor. Alternatively, because KAI1/CD82 physically interacts with b1 integrins and growth factor receptors and downregulates their function, KAI1/CD82 can directly inhibit the promigration signaling at the very upstream. If the complexity of tetraspanin-enriched microdomain constituents is taken into consideration, multiple signaling pathways are susceptible to KAI1/CD82 inhibition. Indeed, we found in this study that KAI1/CD82-induced morphological and cytoskeletal changes could not simply be overridden or bypassed by one or two signaling mechanisms. For example, the signaling originating from b1 integrin, EGFR, c-Met, and CXCR4 promotes cell migration and actin reorganization through Rho small GTPases. Activation of these signaling pathways either alone or together, however, cannot reverse KAI1/CD82-induced morphological and cytoskeletal effects. This observation strongly suggests that KAI1/CD82 acts either on the signaling step at or after the converge point of multiple pathways, e.g., the signaling that immediately leads to actin reorganization, or on the very beginning of multiple signaling pathways, e.g., TEMs, in which integrins and growth factor receptors reside. In either case, KAI1/ CD82 likely acts directly at the plasma membrane where actin reorganization is triggered during cell migration and TEMs are located. The plasma membrane PIP2 is apparently perturbed by KAI1/ CD82 overexpression. Because the PIP2 mAb probes PIP2 after the fixation procedure, it can detect only free PIP2 at the plasma membrane. While GFP-PLCd PH domain fusion not only binds free PIP2 but also likely competes with endogenous PIP2-binding proteins in live cells to occupy PIP2. Thus, the readout of GFPPLCd PH domain fusion more likely reflects the level of free and occupied PIP2 at the plasma membrane. In either case, more PIP2 was found in the plasma membrane of Mock cells. PIP2 links the plasma membrane to actin cytoskeleton by either directly binding and/or activating actin-binding proteins such as b-spectrin, aactinin, vinculin, and ERM proteins or indirectly Gomisin-D inducing the cortical actin polymerization through profilin, cofilin, and NWASP. The level of PIP2 controls the connection of the membrane lipid bilayer to its underlying actin cytoskeleton. The reduced PIP2 at the plasma membrane upon KAI1/CD82 expression, especially at the membrane sites where actin actively undergoes reorganization, likely causes attenuated actin polymerization during cell spreading and migration. Moreover, the composition and distribution of membrane lipid also affect the functional status of Rho GTPases and their effectors because, to be functional, Rho GTPases and their effectors must translocate to the plasma membrane. Because KAI1/CD82 regulates the composition and distribution of PIP2 and other lipids at the plasma membrane, it likely alters the activities of Rho GTPases and their effectors through the translocation step. Interestingly and also surprisingly, cell-cell adhesion significantly alleviates the morphological and cytoskeletal effects of KAI1/ CD82.
earlier, KAI1/CD82 can initiate outside-in signaling. KAI1/CD82-initiated Mepiroxol signaling may intercept the promigration signaling derived from integrin and growth factor receptor. Alternatively, because KAI1/CD82 physically interacts with b1 integrins and growth factor receptors and downregulates their function, KAI1/CD82 can directly inhibit the promigration signaling at the very upstream. If the complexity of tetraspanin-enriched microdomain constituents is taken into consideration, multiple signaling pathways are susceptible to KAI1/CD82 inhibition. Indeed, we found in this study that KAI1/CD82-induced morphological and cytoskeletal changes could not simply be overridden or bypassed by one or two signaling mechanisms. For example, the signaling originating from b1 integrin, EGFR, c-Met, and CXCR4 promotes cell migration and actin reorganization through Rho small GTPases. Activation of these signaling pathways either alone or together, however, cannot reverse KAI1/CD82-induced morphological and cytoskeletal effects. This observation strongly suggests that KAI1/CD82 acts either on the signaling step at or after the converge point of multiple pathways, e.g., the signaling that immediately leads to actin reorganization, or on the very beginning of multiple signaling pathways, e.g., TEMs, in which integrins and growth factor receptors reside. In either case, KAI1/ CD82 likely acts directly at the plasma membrane where actin reorganization is triggered during cell migration and TEMs are located. The plasma membrane PIP2 is apparently perturbed by KAI1/ CD82 overexpression. Because the PIP2 mAb probes PIP2 after the fixation procedure, it can detect only free PIP2 at the plasma membrane. While GFP-PLCd PH domain fusion not only binds free PIP2 but also likely competes with endogenous PIP2-binding proteins in live cells to occupy PIP2. Thus, the readout of GFPPLCd PH domain fusion more likely reflects the level of free and occupied PIP2 at the plasma membrane. In either case, more PIP2 was found in the plasma membrane of Mock cells. PIP2 links the plasma membrane to actin cytoskeleton by either directly binding and/or activating actin-binding proteins such as b-spectrin, aactinin, vinculin, and ERM proteins or indirectly Gomisin-D inducing the cortical actin polymerization through profilin, cofilin, and NWASP. The level of PIP2 controls the connection of the membrane lipid bilayer to its underlying actin cytoskeleton. The reduced PIP2 at the plasma membrane upon KAI1/CD82 expression, especially at the membrane sites where actin actively undergoes reorganization, likely causes attenuated actin polymerization during cell spreading and migration. Moreover, the composition and distribution of membrane lipid also affect the functional status of Rho GTPases and their effectors because, to be functional, Rho GTPases and their effectors must translocate to the plasma membrane. Because KAI1/CD82 regulates the composition and distribution of PIP2 and other lipids at the plasma membrane, it likely alters the activities of Rho GTPases and their effectors through the translocation step. Interestingly and also surprisingly, cell-cell adhesion significantly alleviates the morphological and cytoskeletal effects of KAI1/ CD82.
ROCK activity may not be sensitive enough to report the local diminution of ROCK activity
Leave a reply
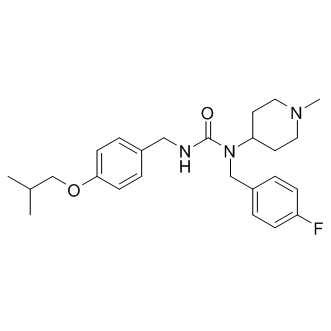 A mouse model of infection with this parasite is available and was used in this study. We used intravital videomicroscopy to analyze the feeding behavior of Anopheles gambiae. We observed the mosquito feeding through the skin of the back of an anesthetized mouse, as described by Petit. The mice were either naive, or had been passively or actively immunized with Anopheles gambiae saliva. The reaction of the skin to Anopheles gambiae blood feedings was followed over time by histological observation. Immunohistochemistry was used to localize the release of saliva and sporozoites, and to follow the course of saliva and sporozoite detection in the skin. Previously reported observations of mosquito bites have mostly concerned bites to frog legs or mouse ears. We used a less specialized model than the ear, which contains cartilage and has a thin skin unlike that covering the rest of the body. We also felt that the observation of Anopheles gambiae mosquitoes biting a mammalian host would be the most appropriate model for obtaining useful data concerning biting and parasite interactions. Our observations are supported by video recordings, still photographs and sections through the tissues of bitten mice. They enabled us to describe in more detail the interaction between Anopheles gambiae and the skin of naive or saliva-sensitized mice. We also used mosquitoes infected with Plasmodium berghei and various tools, including antibodies and fluorescent strains, to study parasite transmission. Videos of the movement of the mosquitoes�� mouthparts within the skin revealed that the tip of the labrum was highly flexible during the probing phase. A large area under the skin was probed, without the mosquito having to withdraw its proboscis or change the point of entry. Similar observations were previously reported by for Aedes aegypti feeding on the webbed skin of frogs. The
A mouse model of infection with this parasite is available and was used in this study. We used intravital videomicroscopy to analyze the feeding behavior of Anopheles gambiae. We observed the mosquito feeding through the skin of the back of an anesthetized mouse, as described by Petit. The mice were either naive, or had been passively or actively immunized with Anopheles gambiae saliva. The reaction of the skin to Anopheles gambiae blood feedings was followed over time by histological observation. Immunohistochemistry was used to localize the release of saliva and sporozoites, and to follow the course of saliva and sporozoite detection in the skin. Previously reported observations of mosquito bites have mostly concerned bites to frog legs or mouse ears. We used a less specialized model than the ear, which contains cartilage and has a thin skin unlike that covering the rest of the body. We also felt that the observation of Anopheles gambiae mosquitoes biting a mammalian host would be the most appropriate model for obtaining useful data concerning biting and parasite interactions. Our observations are supported by video recordings, still photographs and sections through the tissues of bitten mice. They enabled us to describe in more detail the interaction between Anopheles gambiae and the skin of naive or saliva-sensitized mice. We also used mosquitoes infected with Plasmodium berghei and various tools, including antibodies and fluorescent strains, to study parasite transmission. Videos of the movement of the mosquitoes�� mouthparts within the skin revealed that the tip of the labrum was highly flexible during the probing phase. A large area under the skin was probed, without the mosquito having to withdraw its proboscis or change the point of entry. Similar observations were previously reported by for Aedes aegypti feeding on the webbed skin of frogs. The 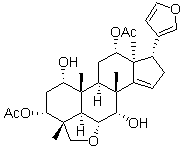 expression can be avoided, possibly allowing for an unbiased view onto changed molecular pathways. Therefore, we decided to run a comparative protein expression study of MPC and MTT cells as a first step to detect proteins that seem to be of importance for an aggressive PHEO/PGL phenotype. Analysis of the differentially expressed proteins focused on changes in energy metabolism related pathways. Expression changes detected in the MPC and MTT cells were then evaluated in human VHL- and SDHB-derived PHEOs/PGLs. Changes related to energy metabolism were further evaluated by measurement of OXPHOS complex activity, ROS production, and expression analysis of additional glycolysis and OXPHOS related genes. The results presented here may lead to better characterization and understanding of the pathogenesis of aggressive PHEOs/PGLs. In the present study, comparative analysis of MPC and MTT cells helped us to identify differentially expressed proteins characteristic of aggressive behavior. We chose this approach to avoid identifying differentially expressed proteins in direct comparisons of SDHB and VHL-PHEOs/PGLs that may be due to tumor location, the
expression can be avoided, possibly allowing for an unbiased view onto changed molecular pathways. Therefore, we decided to run a comparative protein expression study of MPC and MTT cells as a first step to detect proteins that seem to be of importance for an aggressive PHEO/PGL phenotype. Analysis of the differentially expressed proteins focused on changes in energy metabolism related pathways. Expression changes detected in the MPC and MTT cells were then evaluated in human VHL- and SDHB-derived PHEOs/PGLs. Changes related to energy metabolism were further evaluated by measurement of OXPHOS complex activity, ROS production, and expression analysis of additional glycolysis and OXPHOS related genes. The results presented here may lead to better characterization and understanding of the pathogenesis of aggressive PHEOs/PGLs. In the present study, comparative analysis of MPC and MTT cells helped us to identify differentially expressed proteins characteristic of aggressive behavior. We chose this approach to avoid identifying differentially expressed proteins in direct comparisons of SDHB and VHL-PHEOs/PGLs that may be due to tumor location, the