cAMP accumulation, enhanced mitochondrial biogenesis and Ucp1 expression in WAT. WAT in mice with an overexpression of FoxC2 acquired certain BAT-like properties with increased expression levels of PGC1, Ucp1 and cAMP pathway proteins such as b3-adrenoceptor and the Amikacin hydrate protein kinase A alpha regulatory subunit1. Similarly, in mice lacking negative regulators for BAT differentiation like p107, Rb and RIP140, there is a uniform replacement of WAT with BAT and elevated levels of PGC1a and Ucp1. Although many factors and regulatory pathways have been shown to play important roles in maintaining WAT identity, upstream signals or factors that determine the fate of BAT vs WAT and those that initiate the conversion of WAT to BAT remain unclear. Cide proteins including Cidea, Cideb and Fat specific protein 27, share homology with the DNA fragmentation factor DFF40/45 at the N-terminal region. Our previous work demonstrated that Cidea is expressed at high levels in BAT, whereas Cideb is expressed at high levels in liver. Mice with deficiency in both Cidea and Cideb have higher energy expenditure, enhanced insulin sensitivity and are resistance to high-fat-diet -induced obesity and diabetes, suggesting a universal role of Cide proteins in the regulation of energy homeostasis. Fsp27 was originally identified in differentiated ![]() TA1 adipocytes and its expression is regulated by C/ EBP transcription factor. Fsp27 mRNA is detected at high levels in WAT and moderate levels in BAT. Lower levels of Fsp27 mRNA are present in skeletal muscle and lung. In human, it is also detected in heart and many other tissues. Furthermore, Fsp27 protein is detected in the lipid droplet enriched fraction and over-expressed Fsp27 protein was shown to be targeted to lipid droplets and promotes TAG storage. Recently, Fsp27 was also found to be a direct mediator of PPARc-dependent hepatic steatosis. However, its physiological role in regulating adipocyte function and the development of obesity is largely unknown. Here, we observed that Fsp27 protein was detected in both BAT and WAT, was dramatically up-regulated in the WAT and liver of ob/ob mice suggesting that Fsp27 expression is positively correlated with the development of obesity. Fsp27 deficiency results in dramatically reduced WAT depot and the acquisition of a brown fat-like morphology in these WAT, typified by the appearance of Ginsenoside-Ro smaller lipid droplets.
TA1 adipocytes and its expression is regulated by C/ EBP transcription factor. Fsp27 mRNA is detected at high levels in WAT and moderate levels in BAT. Lower levels of Fsp27 mRNA are present in skeletal muscle and lung. In human, it is also detected in heart and many other tissues. Furthermore, Fsp27 protein is detected in the lipid droplet enriched fraction and over-expressed Fsp27 protein was shown to be targeted to lipid droplets and promotes TAG storage. Recently, Fsp27 was also found to be a direct mediator of PPARc-dependent hepatic steatosis. However, its physiological role in regulating adipocyte function and the development of obesity is largely unknown. Here, we observed that Fsp27 protein was detected in both BAT and WAT, was dramatically up-regulated in the WAT and liver of ob/ob mice suggesting that Fsp27 expression is positively correlated with the development of obesity. Fsp27 deficiency results in dramatically reduced WAT depot and the acquisition of a brown fat-like morphology in these WAT, typified by the appearance of Ginsenoside-Ro smaller lipid droplets.
with C/EBPb gene was shown to lead to an increase in the expression of Gsa subunit
Leave a reply
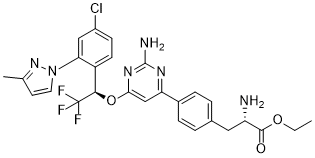 a possible role for ABCA4 in determining pathology in brain in addition to eye, that would be consistent with the association observed with hydrocephalus, in particular when examining associations between mother’s genotype and clinical
a possible role for ABCA4 in determining pathology in brain in addition to eye, that would be consistent with the association observed with hydrocephalus, in particular when examining associations between mother’s genotype and clinical 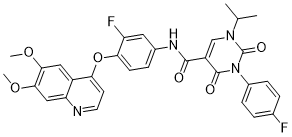 certain Wnt/b-catenin pathway targets to a circumscribed region of the dorsal embryo. It is interesting that effects of Hipk1 on the Wnt/b-catenin pathway depend on an intact kinase domain, and that this kinase activity also appears to negatively regulate the strength of this Hipk1/Tcf3 interaction. This suggests that cooperative functions of Hipk1 and Tcf3 may be regulated by the Hipk1 kinase. In this case Tcf3 or another factor in the same transcriptional complex such as
certain Wnt/b-catenin pathway targets to a circumscribed region of the dorsal embryo. It is interesting that effects of Hipk1 on the Wnt/b-catenin pathway depend on an intact kinase domain, and that this kinase activity also appears to negatively regulate the strength of this Hipk1/Tcf3 interaction. This suggests that cooperative functions of Hipk1 and Tcf3 may be regulated by the Hipk1 kinase. In this case Tcf3 or another factor in the same transcriptional complex such as