In Thailand, B. pseudomallei infections are predominantly manifested as community-acquired septicaemia. Followed by oral antimicrobial therapy comprising of trimethoprim and sulfamethoxazole for 3–6 months. However, despite of adequate antibiotic therapy the pathogen is capable of undergoing latency for decades and relapse in later stages of immune suppressed conditions. No substitutional protecting molecules or human vaccines for the disease currently exist. Therefore, there is a high priority for development of alternative antimicrobial molecules that can efficiently confer passive protection against the disease among the susceptible hosts. To combat the aforementioned challenge, passive antibodymediated protection would be a better alternative as antibodies upon passive administration confer immediate and specific immunity to the susceptible and/or infected host. Additionally, antibodies have minimal toxicity as they are natural in origin. Monoclonal antibodies hold great promise in this view, since the antimicrobial activity conferred by them shall be highly specific. Thus, they are also growing faster as new class of therapeutic and passive protection molecules. Pathogenesis mechanisms of B. pseudomallei are being explored intensively both in vitro and in vivo. As a pathogen, it has several strategies to survive and VX-11e establish pathogenesis in host system. The key factor of its pathogenesis is the ability to survive intracellularly in both phagocytic and non-phagocytic cells. This feature makes the pathogen successful in escaping host immune responses, establish chronic infections, latency and multi-drug resistance. Once invaded into host cells, B. pseudomallei induces cell-to-cell fusion resulting in formation of multinucleated giant cells. This is followed by the induction of apoptosis in infected cells by unknown mechanism. The inherent ability of the pathogen to form biofilm is also likely to be responsible for persistence of infection. B. pseudomallei has demonstrated resistance to antimicrobial drugs when grown in conditions that induce biofilm formation. Development of specific antibodies against the bacterial components involved in adherence and invasion into host cells might possibly prevent the pathogen to establish pathogenesis and in turn confer passive protection in susceptible and infected hosts. Earlier works reported on similar lines implied the promise of structural virulent factors present on the surface of bacteria for the purpose, since location of the antigens plays an important role in determining the effectiveness of the antibodies. Further, the antibodies, when bound to the surface-expressed antigens may enhance phagocytosis or CA-074 Me trigger complement fixation resulting in subsequent killing of internalized bacteria. Targeting a virulence factor that plays a key role in the former mentioned pathogen-mechanisms appears to be promising. However, according to a recent review, passive immunization against B. pseudomallei using mAbs raised against surface expressed molecules including capsular polysaccharide, lipopolysaccharide and exopolysaccharide provide significant but not complete protection in murine models. Also, using mAbs raised against CPS and LPS of B. pseudomallei in different studies have shown partial protection in murine models.
Monthly Archives: July 2019
Two out of three genes coding for gibberellin-20-oxidases involved in the biosynthesis of GAs
Following this observation we were prompted to search for DE genes involved in GA metabolism and signalling. On the other hand, four genes coding for gibberellin-2-oxidases, which inactivate GAs by introducing a hydroxyl at the 2b position, were all upregulated. In rice, under low GA concentrations, the DELLA transcription factor SLR1 represses the GA responses. When GA level increases, the soluble receptor GID1 binds GA, forming the GID1-GA complex. This interacts with SLR1 and leads to its degradation by the 26S proteasome, thus releasing the repressive state of GA responses. In our data, we found that two genes coding for the nuclear gibberellin receptor GID1 were induced in TYLCSV infected plants, and a transcript coding for the GAI protein, a repressor of GA signalling similar to the rice protein SLR1, was repressed. All the above observations suggest a role for GA in plant-TYLCSV interaction, with an increase in GA levels during infection, 6-Maleimidohexanoic acid N-hydroxysuccinimide ester partially counteracted by the induction of the GA-inactivating enzymes gibberellin-2-oxidases. Recent studies revealed a complex crosstalk between GA, ABA and JA signalling mediating plant growth and response to abiotic and biotic stresses. We found that the GO category response to abscisic acid stimulus was overrepresented among up-regulated genes, and the gene coding for the abscisic acid 8’-hydroxylase, a key enzyme for ABA degradation, was down-regulated. These data suggest an increase of ABA level during TYLCSV infection. Actually, in addition to playing a key role in developmental processes and in response to abiotic stresses, ABA has been shown to be involved in the interaction between RNA viruses and their host plants. ABA level was observed to increase in tomato shoots infected by Tomato spotted wilt virus and in tobacco leaves systemically infected by TMV; interestingly, treatment with exogenous ABA improved resistance to TMV infection in tobacco. A first relation between ABA and geminivirus infection was observed in Arabidopsis plants inoculated with the curtovirus, Beet severe curly top virus. ATHB12 and ATHB7 genes, coding for two transcription factors belonging to the homeodomain-leucine zipper family,6-Maleimidocaproic Acid previously shown to be induced by abscisic acid, were also induced in symptomatic BSCTV-infected tissues. Moreover, ATHB12 expression was correlated with several morphological abnormalities such as leaf curling, stunting, and callus-like structures in infected Arabidopsis plants. As far as we know, no other evidence of the ABA involvement in geminivirus infection was found so far. JA has recently received attention in geminivirus research. Although we do not see overrepresentation of genes involved in JA metabolism or in JA response, changes in expression of some individual JA-related transcripts were observed. Salicylic acid is another hormone with an important role in many plant-pathogen interactions and it is known to activate local and systemic defence responses, particularly against biotrophs. Activation of the SA pathway is a typical plant response to RNA viruses and a similar induction of SA-mediated responses was observed in the Arabidopsis-CabLCV infection. Surprisingly, we found nine transcripts, coding for phenylalanine ammonia lyases, key enzymes in the SA biosynthesis, all down-regulated in TYLCSVinfected tissues, thus suggesting a decrease in SA levels in infected plants. Moreover, only two genes coding for proteins belonging to the SA-mediated signalling pathway were regulated during infection, one induced and the other repressed in infected tissues.
More detail whether order transitions in early phases of the change process are a substantial characteristic of psychotherapies
Leave a reply
One alternative explanation could be neural habituation effects as described in social anxiety disorders. It has been shown that stimulation through WZ8040 individualized pictures for the purpose of symptom provocation results in effects distinctly different from neutral, but also from standardized disgust-eliciting pictures. There are other studies that have investigated the results of individualized, system-provoking material, such as pictures from patients’personal environment, verbal descriptions of compulsion-related objects, or tactile stimulation through “dirty” objects. Buchheim et al. used individualized scripts of attachment-relevant themes, which were used to provoke attachment-relevant conflict patterns in repeated fMRI scans during the psychoanalytic process of treating chronically depressed patients. By investigating the role of corticoamygdala circuits in the pathophysiology of OCD, Simon et al. collected a great variety of symptom R428 Axl inhibitor triggers, where patients, in addition to exposure to standardized aversive and neutral pictures, were supposed to provide responses according to the degree of induced obsessive states, anxiety, valence and arousal. Meanwhile, the Maudsley Obsessive-Compulsive Stimuli Set provides a specific and standardized paradigm for symptom-specific provocation of OCD states, which in future could enable the application of individual and standardized pictures to be used in a combined paradigm. Several of the ROIs investigated in this study are part of a comprehensive OCD-related neuronal network and have been identified as change-relevant in a number of outcome studies on the psychotherapy of OCD. For the purpose of quantifying changes, rather large ROIs were selected, which made it impossible to focus on smaller basal ganglia structures, or areas of the amygdala. Other brain regions that have been related to OCD symptoms before did not show reliable responses in the present study, for 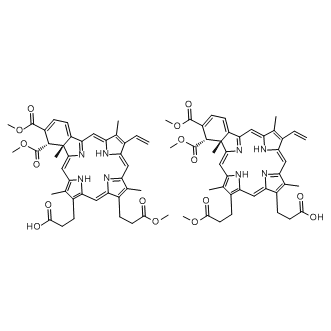 example the ventromedial and the orbitofrontal cortex. The dimension of poor versus good insight could interfere with the results since there seem to exist neuronal correlates of different insight levels. Although patients were classified to the good insight level by their psychiatrists we did not apply a systematic measure of the poor vs. good insight dimension, since studies showed that the degree of insight did not predict the likelihood of response to psychotherapy. This and other possible therapy modulating covariates will be included in a replication study, which offers the possibility to take into consideration the shortcomings of this study. The replication of the study will include a larger sample size and another drug-free diagnostic group in addition to a healthy control group in order to distinguish between OCD-specific activation patterns on the one hand, and diagnosisindependent neuronal mechanisms during therapeutic change processes on the other hand. Furthermore, it would be interesting to perform the scans precisely during periods of low and high dynamic complexity and then compare order transitions with non order transitions as well as phases of critical instability with phases of relative stability. This comparison would be of great theoretical and clinical importance because critical instabilities represent the sensitive phases of therapies. The question would be whether neuronal correlates of instability during therapeutic change processes could be identified. This kind of investigation could focus on the cingulate cortex, which plays an important role as conflict monitoring system. Especially the dorsal part of the ACC is sensitive to ambiguous or conflicting information, is involved in decision making, and its activation is predictive to treatment outcome in depression.
example the ventromedial and the orbitofrontal cortex. The dimension of poor versus good insight could interfere with the results since there seem to exist neuronal correlates of different insight levels. Although patients were classified to the good insight level by their psychiatrists we did not apply a systematic measure of the poor vs. good insight dimension, since studies showed that the degree of insight did not predict the likelihood of response to psychotherapy. This and other possible therapy modulating covariates will be included in a replication study, which offers the possibility to take into consideration the shortcomings of this study. The replication of the study will include a larger sample size and another drug-free diagnostic group in addition to a healthy control group in order to distinguish between OCD-specific activation patterns on the one hand, and diagnosisindependent neuronal mechanisms during therapeutic change processes on the other hand. Furthermore, it would be interesting to perform the scans precisely during periods of low and high dynamic complexity and then compare order transitions with non order transitions as well as phases of critical instability with phases of relative stability. This comparison would be of great theoretical and clinical importance because critical instabilities represent the sensitive phases of therapies. The question would be whether neuronal correlates of instability during therapeutic change processes could be identified. This kind of investigation could focus on the cingulate cortex, which plays an important role as conflict monitoring system. Especially the dorsal part of the ACC is sensitive to ambiguous or conflicting information, is involved in decision making, and its activation is predictive to treatment outcome in depression.
The expression of the myopathy and cataract-causing R120G HspB5 mutant in cells that already
It also represents a cellular model that mimics the early phases of the pathological state before aggresomes of R120G mutated HspB5 accumulate. Expression of wild type HspB5 often correlated with an elongated cellular morphology while expression of the R120G mutant induced the accumulation of membranous ruffles in the leading edges. These effects, which are probably linked to cytoskeleton reorganization, did not change the level of expression of stress markers, such as Hsps. R120G mutant expressing cells were also characterized by perinuclear granules, a phenomenon concomitant with the presence of almost 40% of the cellular content of both HspB1 and HspB5 in the particulate fraction upon cell lysis. Another dominant effect of HspB5 was observed at the level of the native size of HspB1. In presence of type HspB5, HspB1 oligomerization profile was lost and almost 90% of the total cellular amount of this protein interacted with HspB5. HspB5 R120G mutant acted similarly and interacted with HspB1 in a complex of slightly higher molecular mass. Analysis of the resistance to salt-mediated dissociation revealed that HspB1-HspB5 interaction was strengthened by the R120G mutation. This suggests that the interaction was more rigid and less dynamic. The other side of the coin concerned the protective effect of HspB1 towards HspB5. Previous reports mentioned that HspB1 could stabilize mutant HspB5 and attenuate the formation of inclusion bodies induced by expression of R120G HspB5. Using an RNAi approach, we confirmed that, in our cell system, HspB1 attenuates its ubiquitin-proteasome dependent clearance. We report here that cells expressing wild type HspB5 were more oxidoresistant than control cells while those expressing mutant HspB5 displayed a drastically enhanced sensitivity to oxidative stress. HspB5 molecular mechanism that drives oxidoresistance is not yet known. It could result of cytoskeletal protection, since this is a major cellular target rapidly altered in cells exposed to menadione, or modulation of intracellular redox status since HspB1 and HspB5 have been shown to decrease ROS levels and up-regulate the activity of different Lapatinib anti-oxidant enzymes, particularly G6PDH. Moreover, an effect towards HspB1, which gains in chaperone activity once it interacts with HspB5, LY2157299 cannot be excluded. On the other hand, the enhanced sensitivity mediated by the R120G mutation could be a direct consequence of the altered HspB1-HspB5 complex that cannot dissociate in a dynamic and controlled way and the lack of chaperone activity of mutant HspB5. This may lead to a defective recognition of oxidized polypeptides by these chaperones. Analysis of the phosphorylation status pointed to the increased phosphorylation of HspB1 and HspB5 in response to menadione treatment. This study also confirmed the already described hyperphosphorylation of mutant HspB5 in spite of the fact that this polypeptide is in a chimeric complex with HspB1. In that regard, it is interesting to note the R120G mediated enhanced phosphorylation was more intense for the serine sites that are close to the N-terminus; this could disturb hexamer contacts and favor HspB5 oligomer heterogeneity. Two-dimensional immunoblots analysis confirmed the acidic property of the isoforms suggesting that most R120G HspB5 molecules could be phosphorylated. Further analysis following fractionation of the different cell types revealed that the presence of these proteins in the pellet fraction correlated with their sensitivity to oxidative stress. Analysis of the phosphoserine sites of either HspB1 or HspB5 enlightened the different and complex patterns of phosphorylation of these polypeptides, particularly after oxidative stress, which drove a large fraction of these proteins in the particulate fraction. For example, it was first observed 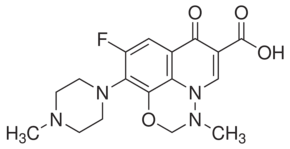 that, in the different untreated cells as well as in menadione-treated Neo.
that, in the different untreated cells as well as in menadione-treated Neo.
Probes for starch showing that the control of the plant circadian clock over its outputs is a process that is constantly changing
Possibly to increase the resonance of the plant’s rhythms with the environmental rhythms. These differences, however, did not impair the sugarcane circadian clock, which suggests that the interactions between the components of the sugarcane circadian clock may also show differences from Arabidopsis. It is worth noting that the presence of common components of the core oscillator between sugarcane and Arabidopsis does not necessarily mean that they will have the same function. It is also possible that there are more differences in the sugarcane circadian clock as we did not try to identify elements of the circadian clock that are present in sugarcane, or grasses, but not in Arabidopsis. One important question is how the circadian clock relays time information to other pathways. We have found 111 transcription factors, of a total of 356 identified in the SUCEST database, regulated by the circadian clock, including ScCCA1 and ScTOC1. Moreover, a large number of probes associated with histone regulation, spliceosome function, RNA degradation and transport are circadian clock regulated. These are mechanisms known to be involved in the control of transcript levels by the circadian clock. Another important regulatory mechanism is the expression of natural antisense transcripts. NATs have been shown to regulate their cognate transcript levels through gene methylation, direct control of transcription, alternative splicing and transcript stability. Widespread antisense transcription has been described in Arabidopsis. In this work, not only have we found AS that were co-regulated with their SS cognates, but AS that had expression levels that did 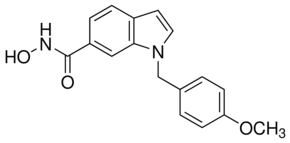 not correlated with their SS cognates, as well. The circadian AS time courses had a tendency to peak towards subjective dawn, while the circadian SS time courses had a tendency to peak towards the subjective middle of the day, a difference that was also observed using tiling arrays in Arabidopsis. AS/SS pairs have been associated with gene plasticity and BIBW2992 EGFR/HER2 inhibitor responsiveness to stimuli, which could also be a strategy to increase the Gefitinib amplitude of circadian rhythms. Most of the circadian AS time courses identified did not have the time courses of their SS cognate considered circadian. These circadian AS may regulate the translation activity of their cognate SS in order to confer circadian behaviour in the protein level. Some AS/SS pairs show small changes in phase, such as violaxanthin deepoxidase and glucose 1-phosphate adenylyltransferase. It is possible that these AS act controlling the expression of their SS cognate, helping to regulate the timing of their expression. The sugarcane circadian transcriptome has shown some properties found in similar works, such as the clustering of transcripts with similar function, the anticipation of daily environmental changes and the temporal compartmentation of metabolic processes. Clustering can be observed in the light harvesting reactions of photosynthesis, in which probes associated with light harvesting complex and photosystem I cluster very tightly between ZT4 and ZT6, when sunlight is usually at its peak. Curiously, probes for synthesis of pigments precede the expression of pigment binding proteins. On the other hand, probes associated with the carbon fixing part of photosynthesis, including RuBisCo activase, tended to cluster between ZT22 and ZT2, in the beginning of the subjective day. Probes for sucrose and starch metabolism are co-ordinated during the day following the timing of the carbon fixing pathways. There is a strong association between the peaks of probes associated with sucrose synthesis, including sucrose phosphatase synthase, with the peaks of transcript for carbon fixation and the upper portion of glycolysis/gluconeogenesis, indicating that one of the destinations of the newly fixed carbon is sucrose.
not correlated with their SS cognates, as well. The circadian AS time courses had a tendency to peak towards subjective dawn, while the circadian SS time courses had a tendency to peak towards the subjective middle of the day, a difference that was also observed using tiling arrays in Arabidopsis. AS/SS pairs have been associated with gene plasticity and BIBW2992 EGFR/HER2 inhibitor responsiveness to stimuli, which could also be a strategy to increase the Gefitinib amplitude of circadian rhythms. Most of the circadian AS time courses identified did not have the time courses of their SS cognate considered circadian. These circadian AS may regulate the translation activity of their cognate SS in order to confer circadian behaviour in the protein level. Some AS/SS pairs show small changes in phase, such as violaxanthin deepoxidase and glucose 1-phosphate adenylyltransferase. It is possible that these AS act controlling the expression of their SS cognate, helping to regulate the timing of their expression. The sugarcane circadian transcriptome has shown some properties found in similar works, such as the clustering of transcripts with similar function, the anticipation of daily environmental changes and the temporal compartmentation of metabolic processes. Clustering can be observed in the light harvesting reactions of photosynthesis, in which probes associated with light harvesting complex and photosystem I cluster very tightly between ZT4 and ZT6, when sunlight is usually at its peak. Curiously, probes for synthesis of pigments precede the expression of pigment binding proteins. On the other hand, probes associated with the carbon fixing part of photosynthesis, including RuBisCo activase, tended to cluster between ZT22 and ZT2, in the beginning of the subjective day. Probes for sucrose and starch metabolism are co-ordinated during the day following the timing of the carbon fixing pathways. There is a strong association between the peaks of probes associated with sucrose synthesis, including sucrose phosphatase synthase, with the peaks of transcript for carbon fixation and the upper portion of glycolysis/gluconeogenesis, indicating that one of the destinations of the newly fixed carbon is sucrose.