We therefore hypothesizethat FIASMAs also penetrate the BBB andare therefore overrepresented in drugs active in the central nervous system. This is supported by the fact that FIASMAs represent different therapeutic classes and different structural classes as evidenced by their ATC-codes. Our results prove that functional inhibition of ASM does not depend on a specific molecular drug class. Instead of specific structural prerequisites, functional inhibition of ASM requires specific physicochemical characteristics of compounds, resulting in high intra-lysosomal concentrations and partitioning of the drug into the inner leaf of the lysosomal membrane. Ceramide and its metabolite sphingosine-1-phosphate have been shown to antagonistically regulate apoptosis, cellular differentiation, proliferation and cell migration. Inhibition of ASM results in anti-apoptotic, proliferative and anti-inflammatory effects. ALK5 Inhibitor II Furthermore, ASM could play a key role in the pathophysiology of depression and in the RAD001 action of antidepressant drugs. Inhibitors of ASM therefore hold promise for a number of new clinical therapies. Several examples are given here: Induction of stroke by experimental ischemia of the brain was shown to correlate with an activation of the ASM and a release of ceramide. Importantly, ASM-deficient mice were protected from tissue damage caused by focal cerebral ischemia. Furthermore, the ethanol-induced neuronal cell death is mediated, at least in part, by activation of ASM and generation of ceramide. In addition, the lethal gastrointestinal syndrome, which limits the efficacy of radiation and chemotherapy, apparently results from apoptotic damage of the endothelial cells of the microvasculature of the small intestine. Genetic inactivation of ASM prevents this toxicity. Infection induced by the human immunodeficiency virus type Iserves as a further example. HIV-1 induces a dramatic depletion of CD4+ T cells in infected individuals, finally leading to AIDS. The massive loss of CD4 T-lymphocytes is assumed to result from apoptosis, probably due to enhanced sphingomyelin breakdown and accumulation of intracellular ceramide. Interestingly, HIV-1 infected long-term nonprogressors have less elevated lymphocyte-associated ceramide than subjects with evolving AIDS, which is paralleled by a lower frequency of apoptotic CD4 and CD8 cells in long-term nonprogressors compared to patients with AIDS. The endotoxic shock syndrome, which is characterized by systemic inflammation, multiple organ damage, circulatory collapse and death, is caused by disseminated endothelial apoptosis sequentially mediated by TNF and ceramide generation. Blocking of this cascade by ASM inhibitors should be preventive and/or therapeutic against the endotoxic shock syndrome. Finally, an imbalance between ASM and acid ceramidase followed by ceramide accumulation has been demonstrated in the hereditary disease cystic fibrosis. In experimental animals, FIASMAs normalize pulmonary ceramide and inflammation. In a clinical pilot study, amitriptyline 6 led to an improved respiratory function in affected patients. Further examples include the treatment of e.g. Alzheimer’s disease, major depression, inflammatory bowel disease, liver cell death and anaemia 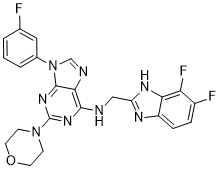 occurring in Wilson disease, and pulmonary edema in acute lung injury. Recently, it has been shown that gilenya, a drug licensed for treatment of multiple sclerosis, acts as a FIASMA. This novel mechanism of action may explain at least in part the beneficial effects of gilenya in multiple sclerosis. AC is a lysosomal enzyme degrading ceramide to sphingosine.
occurring in Wilson disease, and pulmonary edema in acute lung injury. Recently, it has been shown that gilenya, a drug licensed for treatment of multiple sclerosis, acts as a FIASMA. This novel mechanism of action may explain at least in part the beneficial effects of gilenya in multiple sclerosis. AC is a lysosomal enzyme degrading ceramide to sphingosine.
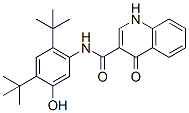
Cationic amphiphilic agents like desipramine chlorpromazine functional inhibition of ASM requires penetration of drugs through biological membranes
Leave a reply
 they do not cross biological membranes via passive diffusion. All compounds were analyzed by their appearance in the ATC system, i.e. substances with multiple effects and different therapeutic indications or different enantiomeric forms were found more than once within the system. In order to determine the distribution of drugs across the ATC system, we used the second level of the 2005 version. Uneven distribution of FIASMAs was tested by a contingency table and one-sided Fisher test. Within this contingency table, the multiple test correctionidentified therapeutic groups with a significantly enriched fraction of FIASMAs. CaM-KIIN is a natural CaMKII inhibitor protein expressed in brain, where CaMKII is also most abundant. The detailed physiological functions of CaMKII inhibition by CaM-KIIN are still unclear. However, precise regulation of CaMKII activity is known to be required for controlling forms of synaptic plasticity underlying higher brain functions such as learning and memory.
they do not cross biological membranes via passive diffusion. All compounds were analyzed by their appearance in the ATC system, i.e. substances with multiple effects and different therapeutic indications or different enantiomeric forms were found more than once within the system. In order to determine the distribution of drugs across the ATC system, we used the second level of the 2005 version. Uneven distribution of FIASMAs was tested by a contingency table and one-sided Fisher test. Within this contingency table, the multiple test correctionidentified therapeutic groups with a significantly enriched fraction of FIASMAs. CaM-KIIN is a natural CaMKII inhibitor protein expressed in brain, where CaMKII is also most abundant. The detailed physiological functions of CaMKII inhibition by CaM-KIIN are still unclear. However, precise regulation of CaMKII activity is known to be required for controlling forms of synaptic plasticity underlying higher brain functions such as learning and memory. and MP are higher than those of RP models solely based on MP. For different combinations fingerprints and MP, the performances of the best RP models were screened according to the C value from different tree depth. For different fingerprint, the best tree depth parameters are different. The best decision tree, with a tree depth of 12 based on FPFP_4 and MP, is shown in Figure 5. The discriminant descriptors include seven MPs and 18 structure fragments. Of the seven MPs chosen by the decision tree, AlogP and logD are properties that describe molecule hydrophobicity, Nrot and nAR describe the molecule’s bulkiness, and MFPSA, MPSA and nHBAcc describe its electrostatic properties or hydrogen binding ability. In other words, the molecular hydrophobicity, size and electrostatic properties are important for mTOR inhibition, which is consistent with previously 3D-QSAR results. Moreover, the eighteen fragments based on FPFP_4 fingerprint also play a key role in discriminating between mTOR inhibitors and non-inhibitors. The na? ��ve Bayesian classifier is an unsupervised learner that does not have a fitting process and tuning parameters, unlike RP method that is sensitive to predefined parameters, e.g., tree depth. The process of Bayesian learning is to
and MP are higher than those of RP models solely based on MP. For different combinations fingerprints and MP, the performances of the best RP models were screened according to the C value from different tree depth. For different fingerprint, the best tree depth parameters are different. The best decision tree, with a tree depth of 12 based on FPFP_4 and MP, is shown in Figure 5. The discriminant descriptors include seven MPs and 18 structure fragments. Of the seven MPs chosen by the decision tree, AlogP and logD are properties that describe molecule hydrophobicity, Nrot and nAR describe the molecule’s bulkiness, and MFPSA, MPSA and nHBAcc describe its electrostatic properties or hydrogen binding ability. In other words, the molecular hydrophobicity, size and electrostatic properties are important for mTOR inhibition, which is consistent with previously 3D-QSAR results. Moreover, the eighteen fragments based on FPFP_4 fingerprint also play a key role in discriminating between mTOR inhibitors and non-inhibitors. The na? ��ve Bayesian classifier is an unsupervised learner that does not have a fitting process and tuning parameters, unlike RP method that is sensitive to predefined parameters, e.g., tree depth. The process of Bayesian learning is to 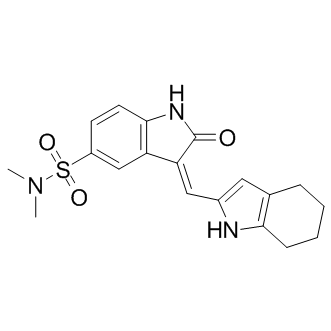 on the inflammatory marker CCL2. Thus, the data presented here are consistent with a function for compound 2 as an inhibitor of NFkB signaling in HRECs. NF-kB has previously been implicated in pathological ocular angiogenesis and we further confirmed that two known NF-kB pathway inhibitors, BAY 11-7082 and CAPE, had antiproliferative effects on HRECs. This assertion of an NF-kB-dependent role for compound 2 can integrate others’ findings regarding the activities of the related compound 1 in other cell types as well. bFGF can act by signaling through phospholipase Cc1, which activates protein kinase C a via diacylglycerol. In turn, PKCa binds and activates IKKa, which phosphorylates and inactivates I-kB.
on the inflammatory marker CCL2. Thus, the data presented here are consistent with a function for compound 2 as an inhibitor of NFkB signaling in HRECs. NF-kB has previously been implicated in pathological ocular angiogenesis and we further confirmed that two known NF-kB pathway inhibitors, BAY 11-7082 and CAPE, had antiproliferative effects on HRECs. This assertion of an NF-kB-dependent role for compound 2 can integrate others’ findings regarding the activities of the related compound 1 in other cell types as well. bFGF can act by signaling through phospholipase Cc1, which activates protein kinase C a via diacylglycerol. In turn, PKCa binds and activates IKKa, which phosphorylates and inactivates I-kB.