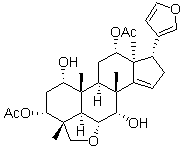
Since the crystal structure of Env trimer is not available, we are not sure if m43 epitope is jointly formed by the CD4bs on gp120 and the N-trimer structure on gp41, or if m43 has dual specificity for one epitope on gp120 and another on gp41, which is very rare. Nevertheless, the m43 epitope is present on functional Env trimer and Pimozide binding of m43 to viral Env prevents virus entry into target cells. Although m43 exhibited modest neutralization activity compared to potent bnmAb VRC01, it neutralized tier 1, 2 and 3 viruses from different clades and four SHIVs tested. M43 showed higher binding to cleavage-incompetent JRFL and Yu2 gp160s than to cleavage-competent JRFL and Yu2 gp160s, which may be attributed to the fact that uncleaved recombinant gp140s were used as antigens during the panning and screening for isolation of m43. Antibody engineering is in progress to improve m43 binding to cleavage-competent JRFL gp160. Improved dual binding to gp120 and gp41 may lead to broader and more potent neutralization activity of the antibody as exemplified by an engineered bispecific anti-HIV-1 antibody that can bind bivalently by virtue of one single chain antibody fragment arm that binds to gp120 and a second arm to the gp41 subunit of gp160. Heterotypic bivalent binding enhanced neutralization compared with the parental antibodies. M43 may represent a new class of bnAbs and its epitope may be used for development of HIV-1 vaccine immunogens. Characterization of the m43 epitope revealed that m43 bound to a conformational epitope that overlaps with the CD4bs on gp120 and the N-trimer structure on gp41. We tried to localize m43 epitope by panning a yeast-displayed Env fragment library against Fab m43, but no enrichment was observed. CD4bs mAbs strongly competed with m43 for binding to recombinant gp140s and membrane-associated functional Env trimers, suggesting that m43 epitope overlaps with the CD4bs. The coreceptor binding site may not be involved in m43 binding as evidenced by lack of correlation between coreceptor density on target cells and m43 neutralization activity. The modest competition of CD4i mAbs with m43 for binding to the Env may be due to a steric hindrance. The effect of sCD4 on m43 binding to recombinant gp140s and membrane-associated Envs is different. sCD4 weakly inhibited m43 binding to recombinant gp14089.6, while sCD4 enhanced m43 binding to cleavage-competent gp160JRFL on 293T cells. Mating produces ![]() a uniform number of males and hermaphrodites because the 3,4,5-Trimethoxyphenylacetic acid larger male sperm replace hermaphrodite sperm during fertilization, which results in an equal segregation of sex chromosomes. Although males are smaller than hermaphrodites, they contain more somatic cells. Most physical differences between the sexes occur during larval development because males and hermaphrodites are nearly identical upon hatching. Beginning at the L4 larval stage, the two sexes are distinguishable using stereomicroscopy. At this stage, the blunt-ended tail of the male starts to develop. The postembryonic development of sex-specific cell lineages and differentiated cells has been previously examined. Furthermore, the anatomy of the male differs from that of the hermaphrodite by having one gonadal arm and having specialized muscles and neurons. Adult hermaphrodites have a peaked tail, a vulva, a uterus and two gonadal arms, which produce sperm and oocytes. The pharynx, excretory system and main body muscles do not exhibit sexually dimorphic characteristics. Due to the existence of sex-specific neurons and sex-related differences in the core nervous system, C. elegans exhibit a sexually dimorphic nervous system.
a uniform number of males and hermaphrodites because the 3,4,5-Trimethoxyphenylacetic acid larger male sperm replace hermaphrodite sperm during fertilization, which results in an equal segregation of sex chromosomes. Although males are smaller than hermaphrodites, they contain more somatic cells. Most physical differences between the sexes occur during larval development because males and hermaphrodites are nearly identical upon hatching. Beginning at the L4 larval stage, the two sexes are distinguishable using stereomicroscopy. At this stage, the blunt-ended tail of the male starts to develop. The postembryonic development of sex-specific cell lineages and differentiated cells has been previously examined. Furthermore, the anatomy of the male differs from that of the hermaphrodite by having one gonadal arm and having specialized muscles and neurons. Adult hermaphrodites have a peaked tail, a vulva, a uterus and two gonadal arms, which produce sperm and oocytes. The pharynx, excretory system and main body muscles do not exhibit sexually dimorphic characteristics. Due to the existence of sex-specific neurons and sex-related differences in the core nervous system, C. elegans exhibit a sexually dimorphic nervous system.
We identified and extensively characterized a novel cross-reactive that recognizes a new conserved neutralizing epitope shared
Leave a reply
 expression can be avoided, possibly allowing for an unbiased view onto changed molecular pathways. Therefore, we decided to run a comparative protein expression study of MPC and MTT cells as a first step to detect proteins that seem to be of importance for an aggressive PHEO/PGL phenotype. Analysis of the differentially expressed proteins focused on changes in energy metabolism related pathways. Expression changes detected in the MPC and MTT cells were then evaluated in human VHL- and SDHB-derived PHEOs/PGLs. Changes related to energy metabolism were further evaluated by measurement of OXPHOS complex activity, ROS production, and expression analysis of additional glycolysis and OXPHOS related genes. The results presented here may lead to better characterization and understanding of the pathogenesis of aggressive PHEOs/PGLs. In the present study, comparative analysis of MPC and MTT cells helped us to identify differentially expressed proteins characteristic of aggressive behavior. We chose this approach to avoid identifying differentially expressed proteins in direct comparisons of SDHB and VHL-PHEOs/PGLs that may be due to tumor location, the
expression can be avoided, possibly allowing for an unbiased view onto changed molecular pathways. Therefore, we decided to run a comparative protein expression study of MPC and MTT cells as a first step to detect proteins that seem to be of importance for an aggressive PHEO/PGL phenotype. Analysis of the differentially expressed proteins focused on changes in energy metabolism related pathways. Expression changes detected in the MPC and MTT cells were then evaluated in human VHL- and SDHB-derived PHEOs/PGLs. Changes related to energy metabolism were further evaluated by measurement of OXPHOS complex activity, ROS production, and expression analysis of additional glycolysis and OXPHOS related genes. The results presented here may lead to better characterization and understanding of the pathogenesis of aggressive PHEOs/PGLs. In the present study, comparative analysis of MPC and MTT cells helped us to identify differentially expressed proteins characteristic of aggressive behavior. We chose this approach to avoid identifying differentially expressed proteins in direct comparisons of SDHB and VHL-PHEOs/PGLs that may be due to tumor location, the 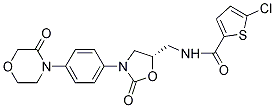 erythrocyte-binding domain. It is possible the variable expression of PfRh4 is used a means of immune evasion, rather than immune evasion via the evolution of polymorphisms. The limited polymorphism in PfRh4 may make it attractive as a vaccine candidate, especially when partnered with other antigens, such as EBA175, to block the use of alternate invasion pathways. PfRh5 also has limited polymorphisms, and a recent report suggested that there was minimal acquisition of IgG to recombinant PfRh5 in a population of malaria-exposed Kenyans. One study reported that expression of the pfrh5 gene was variable among P. falciparum isolates in The Gambia, as seen for other pfrh genes, whereas other data suggests PfRh5 protein is expressed by all laboratory-adapted isolates. Our preliminary studies suggest that the prevalence of antibodies to recombinant PfRh5 is high in our PNG study population. Further studies to understand the expression of PfRh ligands and the acquisition of antibodies to them will be valuable for understanding the importance of PfRh proteins as targets of immunity and their potential for vaccine development. Understanding the targets of acquired human immunity is valuable for several reasons. One of the important criteria for objectively prioritizing antigens for vaccine development against malaria, as with other infectious pathogens, is the demonstration that immune responses to that antigen are associated with protection from malaria.
erythrocyte-binding domain. It is possible the variable expression of PfRh4 is used a means of immune evasion, rather than immune evasion via the evolution of polymorphisms. The limited polymorphism in PfRh4 may make it attractive as a vaccine candidate, especially when partnered with other antigens, such as EBA175, to block the use of alternate invasion pathways. PfRh5 also has limited polymorphisms, and a recent report suggested that there was minimal acquisition of IgG to recombinant PfRh5 in a population of malaria-exposed Kenyans. One study reported that expression of the pfrh5 gene was variable among P. falciparum isolates in The Gambia, as seen for other pfrh genes, whereas other data suggests PfRh5 protein is expressed by all laboratory-adapted isolates. Our preliminary studies suggest that the prevalence of antibodies to recombinant PfRh5 is high in our PNG study population. Further studies to understand the expression of PfRh ligands and the acquisition of antibodies to them will be valuable for understanding the importance of PfRh proteins as targets of immunity and their potential for vaccine development. Understanding the targets of acquired human immunity is valuable for several reasons. One of the important criteria for objectively prioritizing antigens for vaccine development against malaria, as with other infectious pathogens, is the demonstration that immune responses to that antigen are associated with protection from malaria. suboptimal at even high concentrations of anti-EV antibody. Therefore, understanding the mechanism by which anti-EV antibodies provide protection has been of interest. Recent mouse studies have elucidated that an IgG2a isotype monoclonal antibody against the B5 protein called B126 can neutralize EV in the presence of complement and utilizes C’ to partially mediate protection in vivo. This evidence suggests that antibody against EV would be more effective if it was of an isotype that mediated effector functions such as activation of C’ and/or Fc receptor dependent activity. Previous studies of antibody responses to protein vaccination found that formulations that included adjuvants that produced higher titers of IgG2a antibody in mice and IgG1 antibody in non-human primates were more effective at mediating protection than vaccines formulated without these adjuvants. This suggests that antibody with specific Fc activities might be beneficial for protection. By utilizing a high PFU luciferase reporter EV neutralization assay, we find that polyclonal antibody responses against the EV proteins A33 and B5 utilize C’ to neutralize virus in vitro, though in mechanistically different ways. These findings shed light on how differing viral proteins dictate the requirements for the host to neutralize incoming virus with C’. Additionally, we show that antibody against B5 utilizes C’ and FcRs to protect mice from lethal VACV challenge. These findings add to our understanding of how antibody can protect against orthopoxvirus disease and highlights the importance of understanding antibody effector functions necessary for protection to aid in the rational design of anti-viral vaccines and therapeutic antibodies. Proteins used in the vaccine formulations were purified recombinant baculovirus-expressed proteins that were previously described. Protein vaccines were prepared and used as described previously. Similar results were obtained when rabbit pAb against A33 and B5 was used. These results are consistent with the previously described findings that the mechanism of C’-mediated neutralization for A33 and B5 differ, with anti-A33 antibody relying on virolysis and anti-B5 antibody able to neutralize by opsonization. Benhnia hypothesized that antibody alone was unable to fulfill the basic occupancy model for EV neutralization because of the amount of B5 protein on the EV surface and that antibodyinduced C’ coating of the EV membrane allowed for the occupancy model to succeed.
suboptimal at even high concentrations of anti-EV antibody. Therefore, understanding the mechanism by which anti-EV antibodies provide protection has been of interest. Recent mouse studies have elucidated that an IgG2a isotype monoclonal antibody against the B5 protein called B126 can neutralize EV in the presence of complement and utilizes C’ to partially mediate protection in vivo. This evidence suggests that antibody against EV would be more effective if it was of an isotype that mediated effector functions such as activation of C’ and/or Fc receptor dependent activity. Previous studies of antibody responses to protein vaccination found that formulations that included adjuvants that produced higher titers of IgG2a antibody in mice and IgG1 antibody in non-human primates were more effective at mediating protection than vaccines formulated without these adjuvants. This suggests that antibody with specific Fc activities might be beneficial for protection. By utilizing a high PFU luciferase reporter EV neutralization assay, we find that polyclonal antibody responses against the EV proteins A33 and B5 utilize C’ to neutralize virus in vitro, though in mechanistically different ways. These findings shed light on how differing viral proteins dictate the requirements for the host to neutralize incoming virus with C’. Additionally, we show that antibody against B5 utilizes C’ and FcRs to protect mice from lethal VACV challenge. These findings add to our understanding of how antibody can protect against orthopoxvirus disease and highlights the importance of understanding antibody effector functions necessary for protection to aid in the rational design of anti-viral vaccines and therapeutic antibodies. Proteins used in the vaccine formulations were purified recombinant baculovirus-expressed proteins that were previously described. Protein vaccines were prepared and used as described previously. Similar results were obtained when rabbit pAb against A33 and B5 was used. These results are consistent with the previously described findings that the mechanism of C’-mediated neutralization for A33 and B5 differ, with anti-A33 antibody relying on virolysis and anti-B5 antibody able to neutralize by opsonization. Benhnia hypothesized that antibody alone was unable to fulfill the basic occupancy model for EV neutralization because of the amount of B5 protein on the EV surface and that antibodyinduced C’ coating of the EV membrane allowed for the occupancy model to succeed.